スケトウダラ太平洋系群は、数年に一度の割合で発生する卓越年級群(特に加入量が多い年の群)によって資源水準が維持されると考えられています。そのため、卓越年級群の発生機構を明らかにすることがスケトウダラ太平洋系群の資源変動要因を解明し、的確な資源評価・資源管理方策を行う上での重要な要素になっています。
そこで本課題では、通常年と卓越年級群の発生年における稚仔魚期の海流による輸送状況の違いを検討すると共に、成長差に伴う捕食減耗の違いを基に卓越年級群の発生要因を明らかにし、スケトウダラ太平洋系群の資源変動機構を解明することを目指しています。
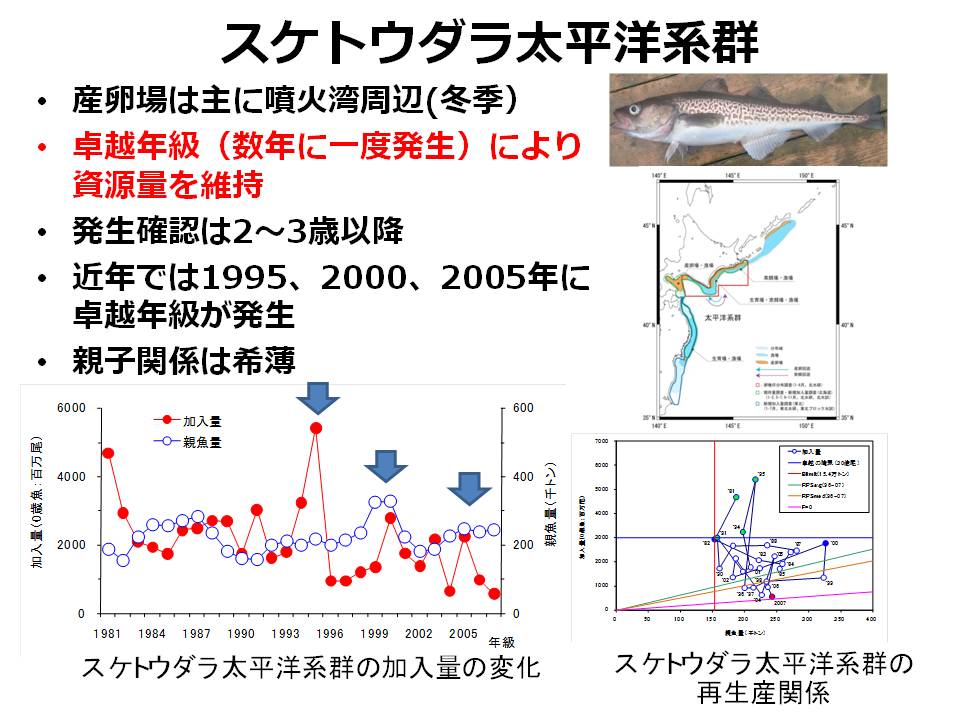
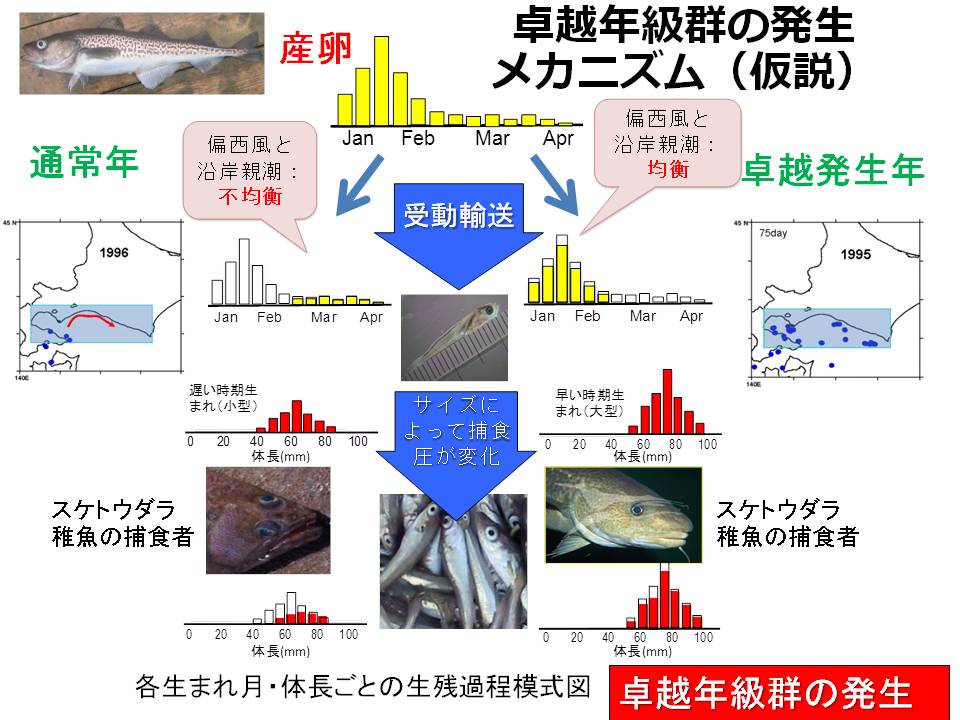

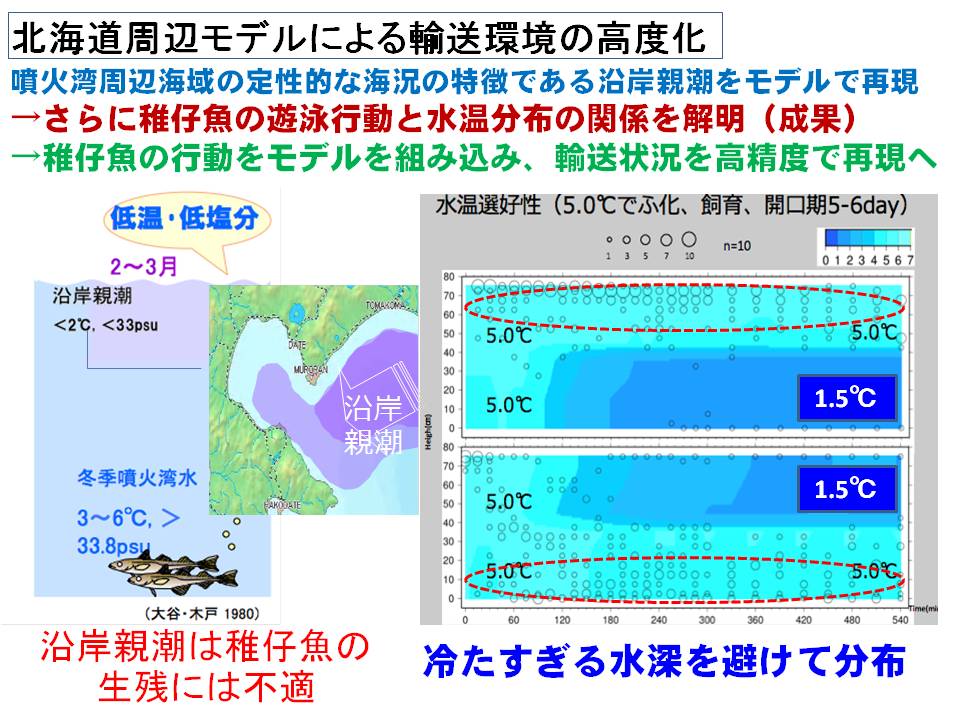
スケトウダラ太平洋系群では、冬季に産卵された卵稚仔魚のうち、噴火湾に春季まで滞留した仔稚魚が、スケトウダラ太平洋系群の加入に貢献すると考えられています。また、近年の稚魚の成長履歴と加入量を調べると、春季以降に噴火湾から道東養育場まで回遊する間の成長・生残も、加入量に影響すると考えられるようになってきました。そこで、稚仔魚の分布特性(沿岸親潮との関連、分布水深)を明らかにするとともに、噴火湾から道東海域の養育場までの輸送モデルを高度化することで、輸送環境が及ぼすスケトウダラ稚魚の成長・生残への影響を明らかにすることを試みています。
スケトウダラ太平洋系群が被食の割合によって変動し、加入量に大きな変動をもたらすのは当歳の5月以降~翌春までと考えられます。そこで、道東養育場に着底する以前の遊泳生活期(主に日高湾内)における被食の影響把握が重要です。これまでの調査結果では、特にソウハチやスケトウダラ(共食い)による影響が大きいことが明らかになってきました。特に、カタクチイワシがいない場合、これらの魚種によってスケトウダラ幼稚魚が捕食される割合が高く、影響が大きいことが明らかになってきました。今後、捕食者量の変動との関連を把握するとともに、環境変動がトップダウン・コントロールに及ぼす変化を定量化していきます。


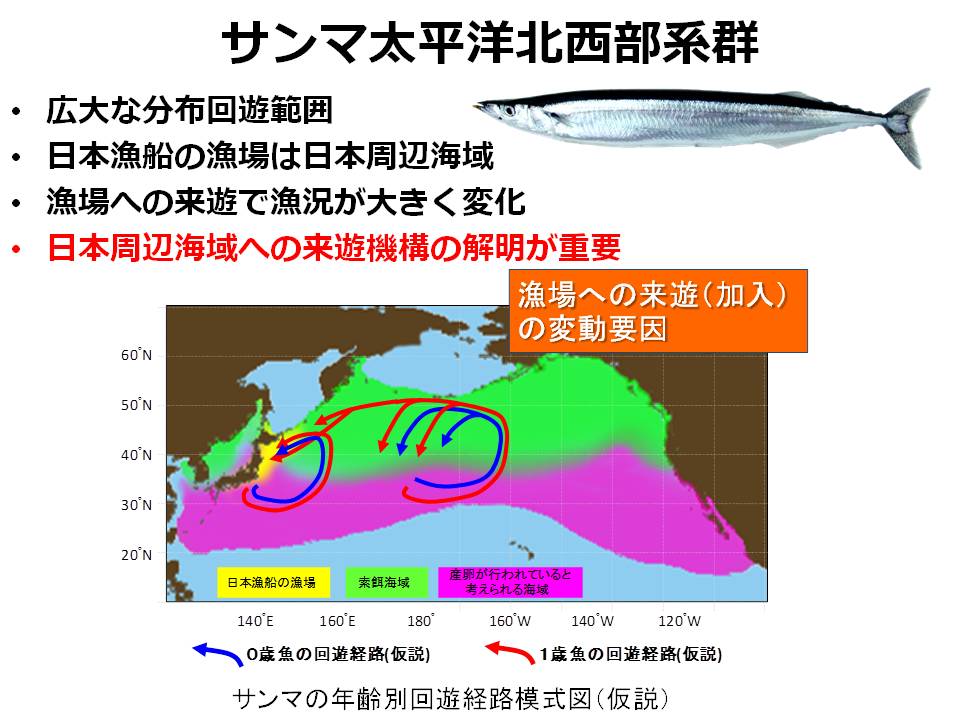
サンマは太平洋北西部に広く分布し、膨大な資源量があります。しかし、日本漁船が生鮮として水揚げ可能なサンマは、日本周辺海域に来遊した群のみとなっています。そのため、サンマが日本周辺域にどのように来遊してくるかを明らかにすることが我が国におけるサンマの安定供給および適確な資源管理を行う上での重要な要素になっています。
そこで本課題では、分布海域によるサンマの成長の違い、および年齢(サンマの寿命は2歳)による回遊パターンの違いを基に日本周辺域に来遊してくるサンマの来遊要因を明らかにし、日本周辺海域への加入量変動機構を明らかにすることを目指しています。
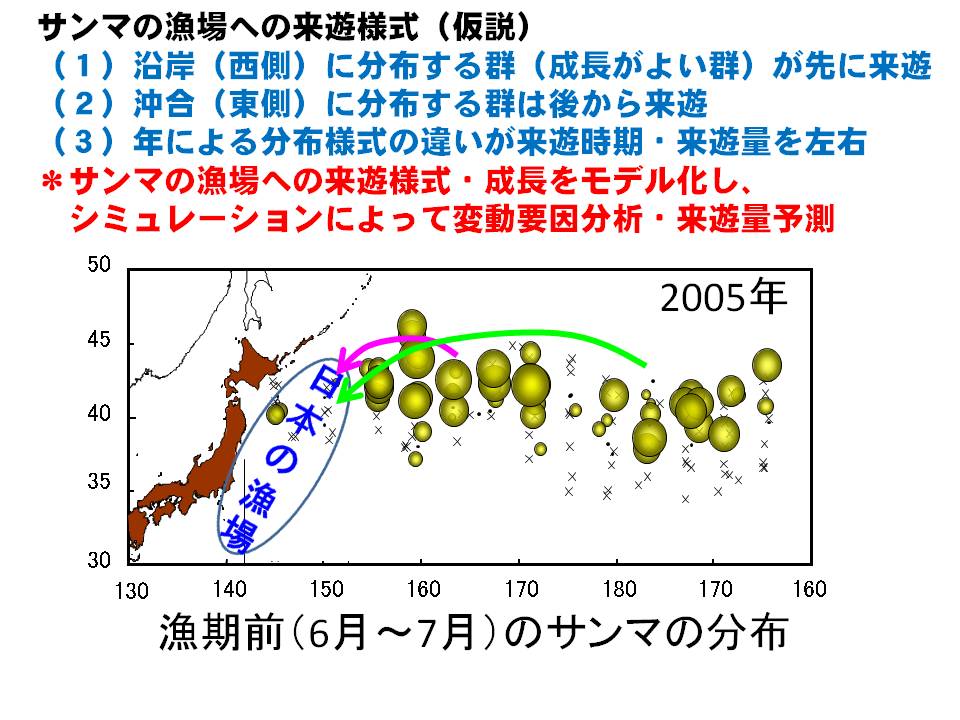
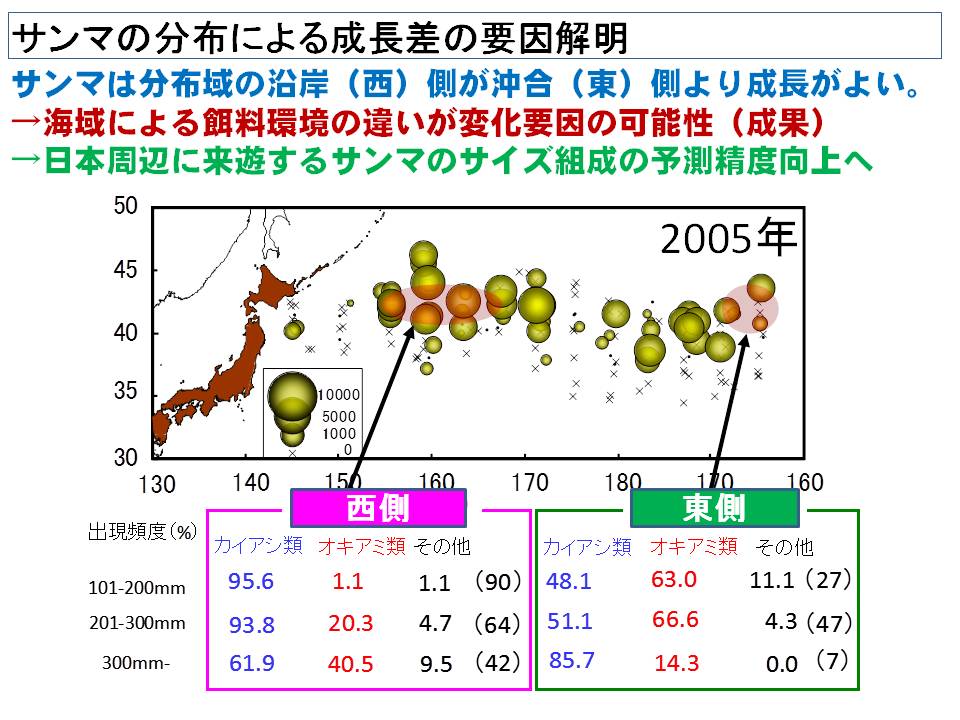
6月~7月に採集されたサンマを調べると、日本に近い海域(東経域)に分布するサンマは成長が早く、日本のはるか沖(西経域)に分布するサンマは成長が遅いことが分かってきました。この成長の違いは、餌の量と質が影響している可能性がこれまでの調査結果から示されています。
サンマの重要な餌はカイアシ類とオキアミ類でしたが、成長の速い東経域では小さいころ(200mm以下)からカイアシ類を食べていた個体が多かったのに対し、西経域ではオキアミ類を食べていた個体が多いことが分かりました。また、成長に伴って東経域ではオキアミ類の利用率が、西経域ではカイアシ類の利用率が高くなっていました。今後はさらに広い海域で採集されたサンマの餌生物を調べてこの傾向を確認するとともに、成長との関連をさらに調査していく予定です。
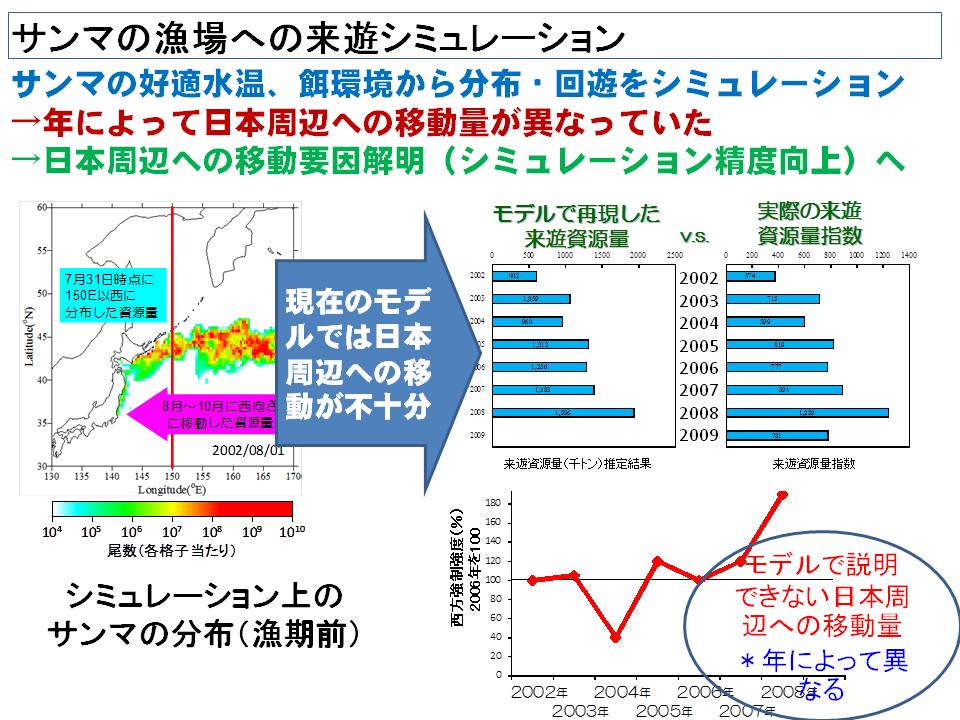
6月~7月に日本のはるか沖(西経域)に分布していたサンマは、その後、日本の沿岸に向かって移動し、サンマ漁が始まる8月~9月には日本周辺の漁場に到達します。しかし、この日本周辺海域に向かう西方向の移動量は年によって変わることが明らかとなりました。6月~7月(分布調査時期)のサンマの分布海域と、8月~9月(漁期開始時期)の分布海域を比較すると、西向きの移動量は2004年が最も低く、2008年が最も高いと計算されました。つまり、多くのサンマが西に向かう年とそうではない年があり、その違いが日本周辺海域へのサンマの来遊量を左右する可能性が示されました。
現在、サンマの西経域から日本周辺海域への移動要因は明らかにされてなく、回遊モデルを用いても来遊量を再現するには至っていません。今後は、どのような海洋の環境の変化が西側へのサンマの移動を促しているか(あるいは阻害しているか)を明らかにし、日本沿岸域へのサンマの来遊機構を明らかにしていく予定です。

マイワシ・マサバ太平洋系群は、海洋環境の中長期的(10年~数10年周期)変化と同調して資源変動する特徴があり、とくにマイワシで顕著です。そのため、両系群の資源評価・管理方策を検討する上で、加入量の変動要因を明らかにすることが重要な課題になっています。また、現在、両系群とも資源水準が中位水準に回復し、毎年の加入量を早期に高い精度で把握することが、若齢時から漁獲対象となる両系群の資源評価およびTAC(漁獲可能量)の算定精度を向上させる鍵になっています。

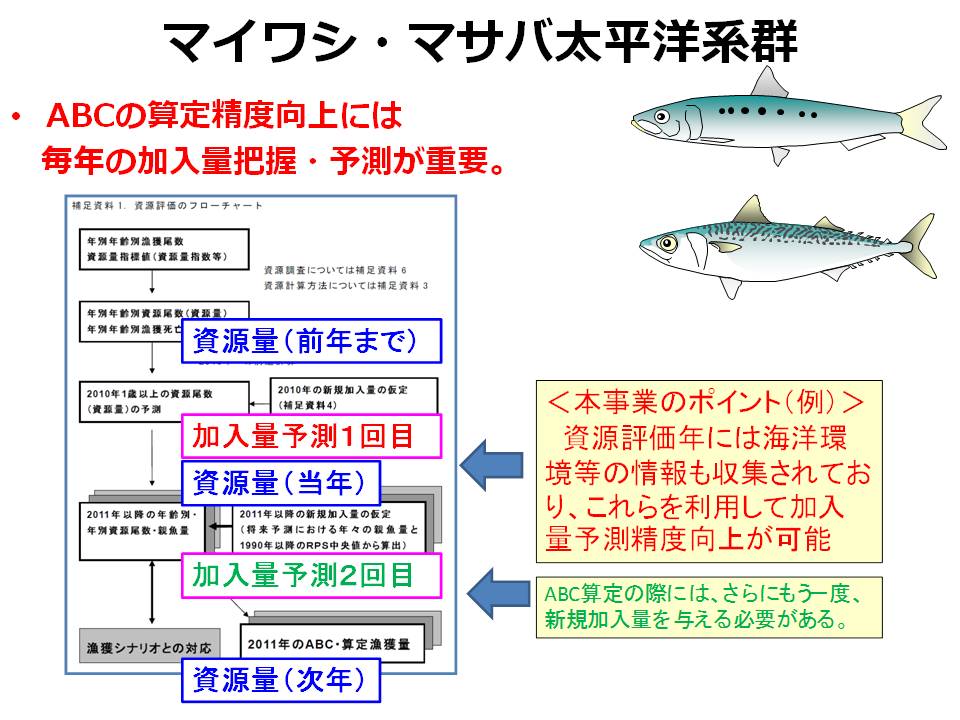
そこで本課題では、年による成長や生残の違いと環境要因との関連を調べることで、「いつ、どのような海洋環境の変化によって加入量が変動するか」、を明らかにすることを目的としています。特に、加入量が決まる時期、海域、発育段階、および決定メカニズムを現場観測、飼育実験、ならびに卵・仔稚幼魚の回遊・成長モデルを用いて解析する計画です。また、現場観測による加入量の早期把握とその精度の向上に取り組むとともに、海洋環境予測に基づく加入量予測モデルの構築も目指しています。
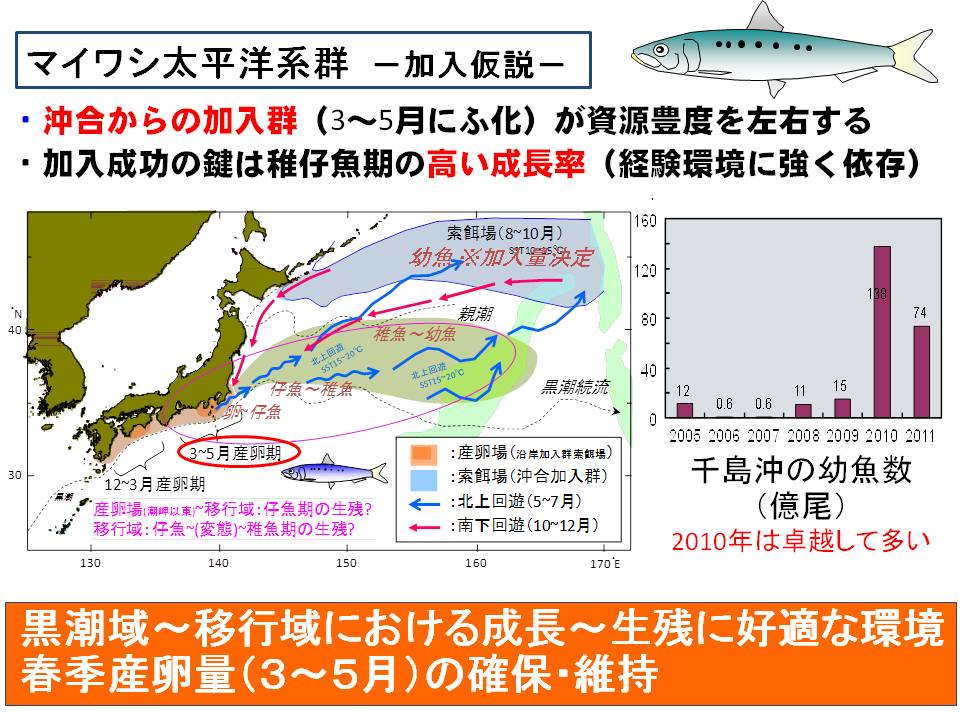

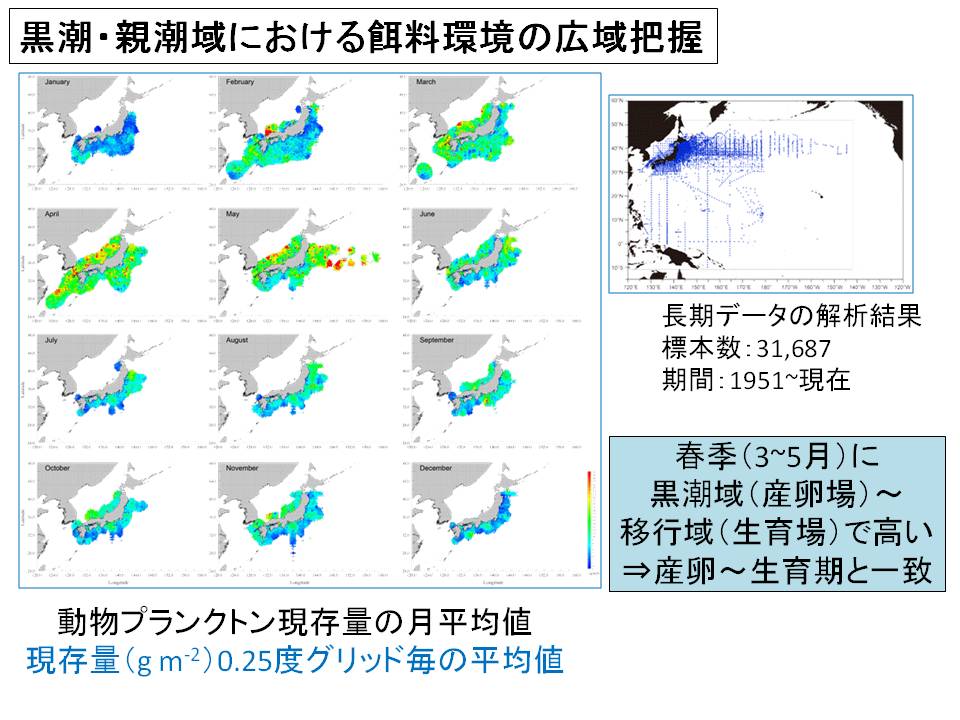
マイワシ太平洋系群は、沖合加入群(4月を中心とする春季にふ化して千島沖の黒潮・親潮移行域で成長、加入する群)の多寡によって資源水準が左右されることがわかってきました。沖合加入群の生残率は成長率と深く関係することが指摘されており、特に加入量が多かった2010年級群は稚仔魚期に成長率の高い個体の割合が例年より高かったことがわかっています。つまり、黒潮・親潮移行域の稚仔魚期の成長率がマイワシ太平洋系群の加入量を左右する要因と考えられます。また、飼育下におけるマイワシ親魚の産卵誘導に初めて成功し、飼育実験用の卵、稚仔魚を得るための飼育実験系が確立しつつあります。今後、飼育実験による稚仔魚期の成長、生残と環境要因との関係を解明し、加入量予測手法の開発を進めていく予定です。
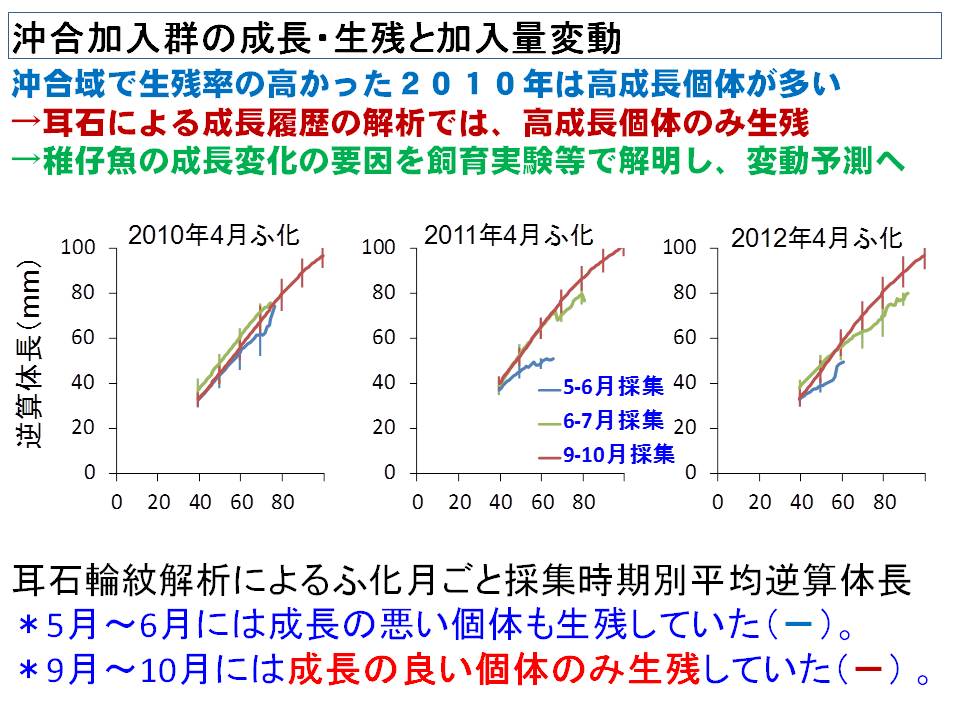

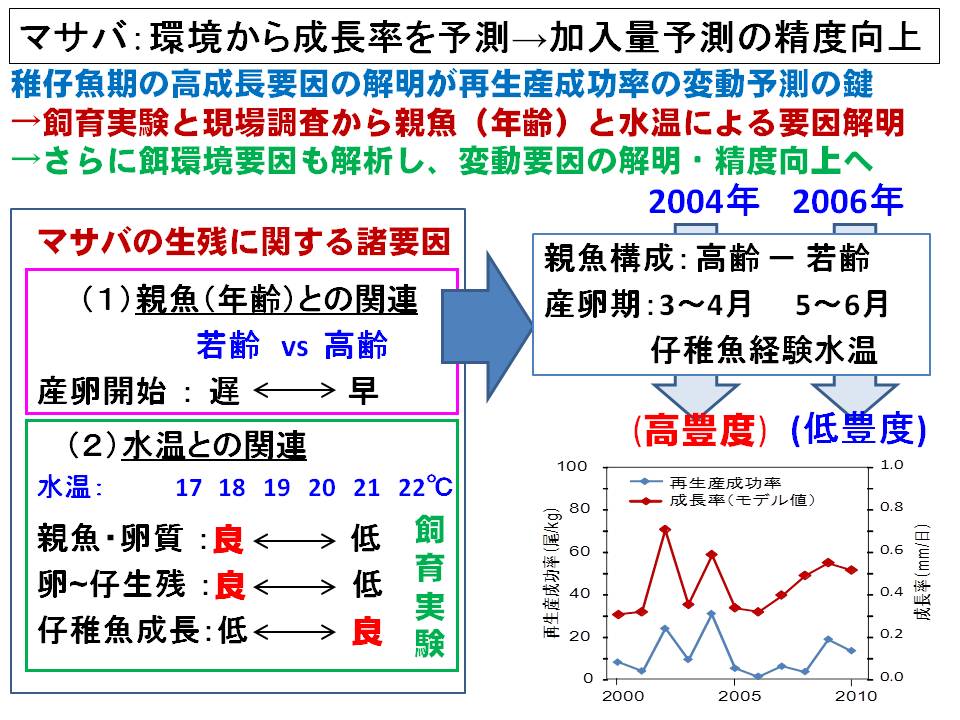
マサバ太平洋系群の毎年の加入量は主産卵期である4月生まれの生き残りによって決まることがわかってきました。そして4月生まれの稚仔魚期の成長率が高いと加入量(生き残り)が高くなる関係が調査結果から把握されると共に、飼育実験によって稚仔魚期の成長に与える水温の影響が明らかになってきました。さらに、親魚の状態(産卵経験の有無、栄養状態、経験環境等)が産み出される卵、仔魚の生残に与える影響(母性効果)が明らかになってきています。
これらの結果から、年々の稚仔魚期の経験水温をもとに成長、生残、加入量を予測するモデルを作成し、さらに精度向上のため、餌量や母性効果などの環境要因を加えるなどの改良を進めています。

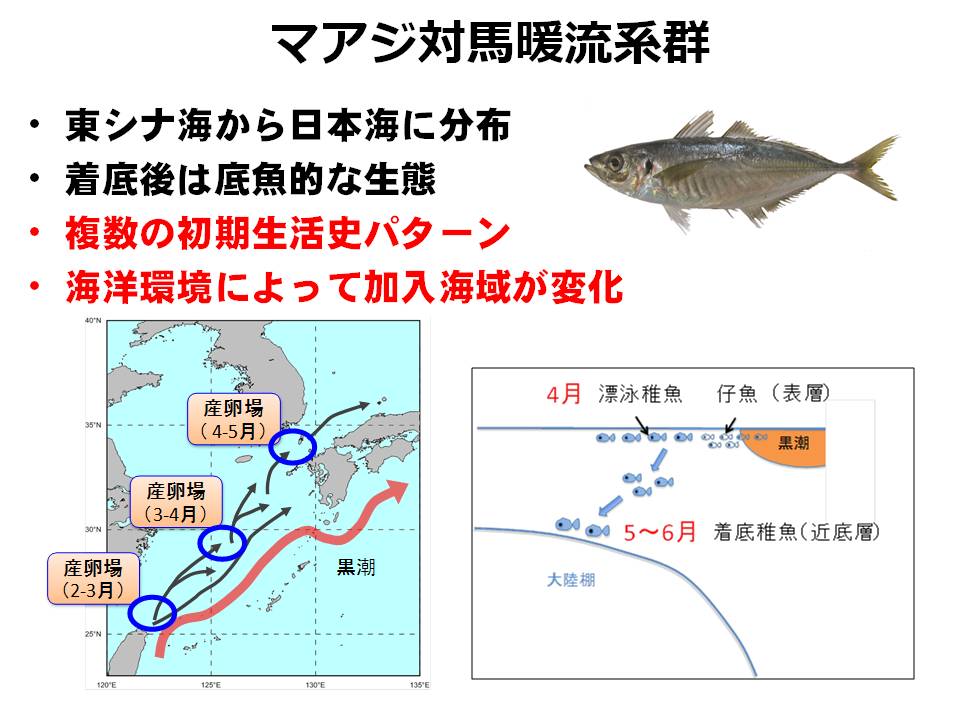
マアジ対馬暖流系群は東シナ海~日本海に広く分布しています。産卵期間も長く、幼稚魚は複数の生活史パターン(表層を泳ぐ、底層に留まる ← 成長速度によって異なる)を持つことから、産卵時期・産卵海域と年々の海洋環境の変化によって加入する海域、加入量が変化する特徴があります。そのため、海洋環境の変化によってどこの海域にどれくらい加入するかを明らかにすることがマアジ対馬暖流系群の資源評価およびTAC(漁獲可能量)の算定精度を向上させる上での重要な課題となっています。
そこで本課題では、餌環境による成長の違いを明らかにすると共に、生まれた時期・海域と海洋環境(餌環境と稚仔魚の流され方)からどの海域(東シナ海または日本海)に多く加入するかをシミュレーションによって早期に把握する手法の開発を行っています。
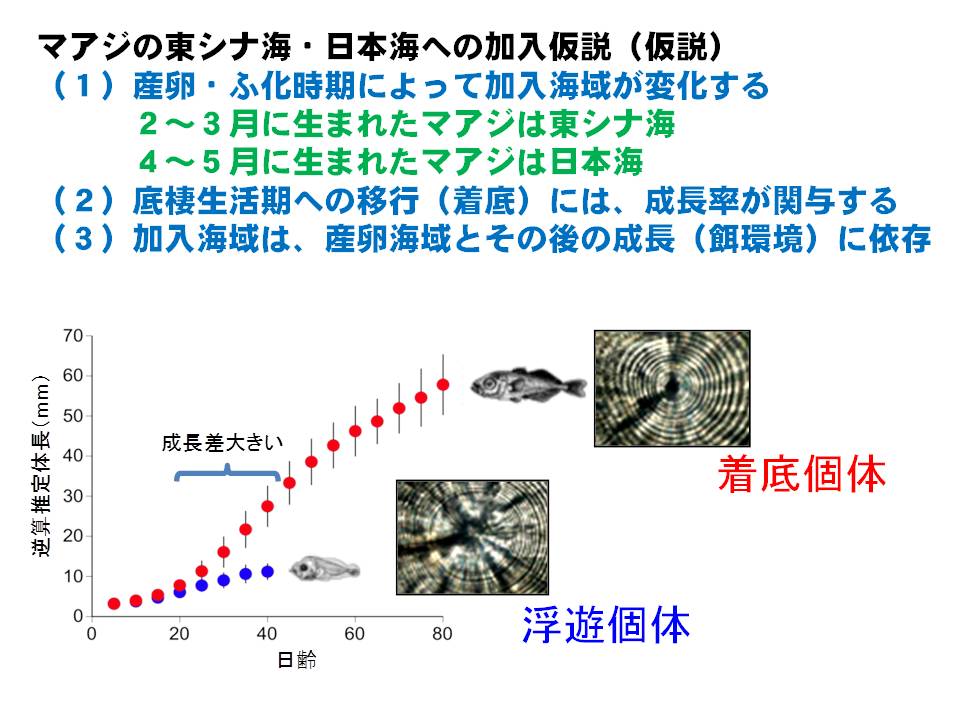
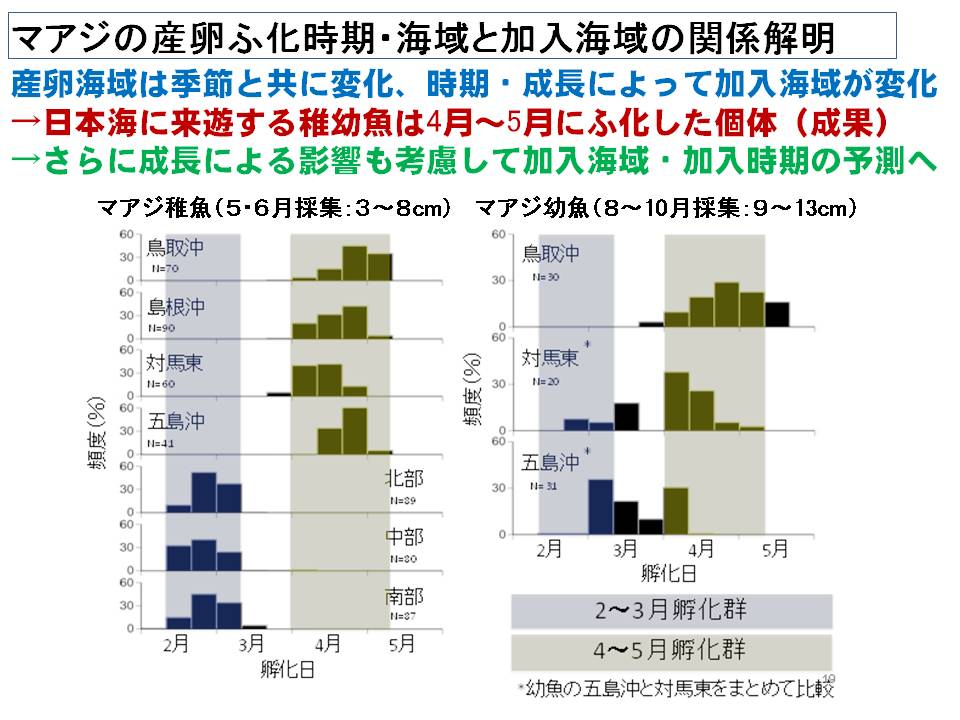
まず、各海域で採集されるマアジの稚魚・幼魚のふ化時期を耳石の日輪解析をもとに調べました。その結果、東シナ海に加入するマアジは主に2月~3月にふ化したマアジが多いのに対し、日本海に加入するマアジは4月~5月にふ化したマアジが多いことがわかりました。また、日本海で加入するマアジは東シナ海に加入するマアジに比べて仔稚魚期の成長が遅いことが明らかになりました。その要因として、日本海では主要な餌であるカイアシ類の密度が東シナ海よりも低い傾向であることが推察されました。
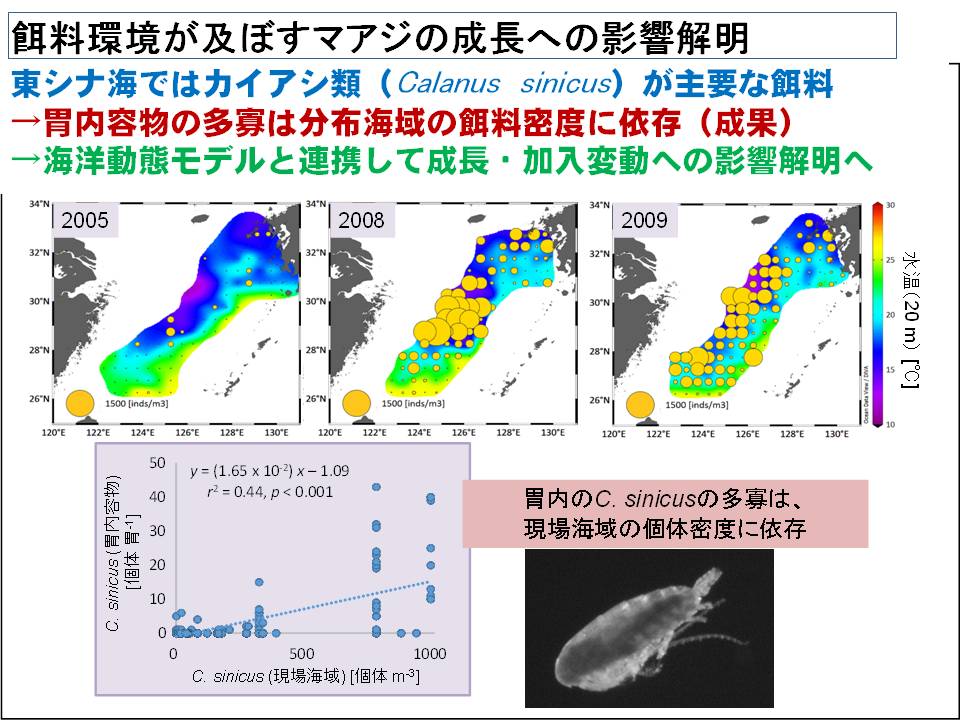
2月~3月のマアジ産卵場は台湾の北付近にあると推定されています。現在、この海域からのマアジ幼稚魚の流され方、および成長について、海洋環境のシミュレーションをもとに検討を進め、加入海域を推定する手法の開発を行っています。なお、海流に流されている間、稚仔魚の成長・生残に適切と判断される水温帯(18.5℃~23.5)の面積と東シナ海のマアジ稚魚の分布密度は深く関係していることも明らかになってきました。今後、海洋環境のシミュレーションを改良しながら研究を進め、東シナ海および日本海に加入するマアジの変動要因を明らかにしていく予定です。

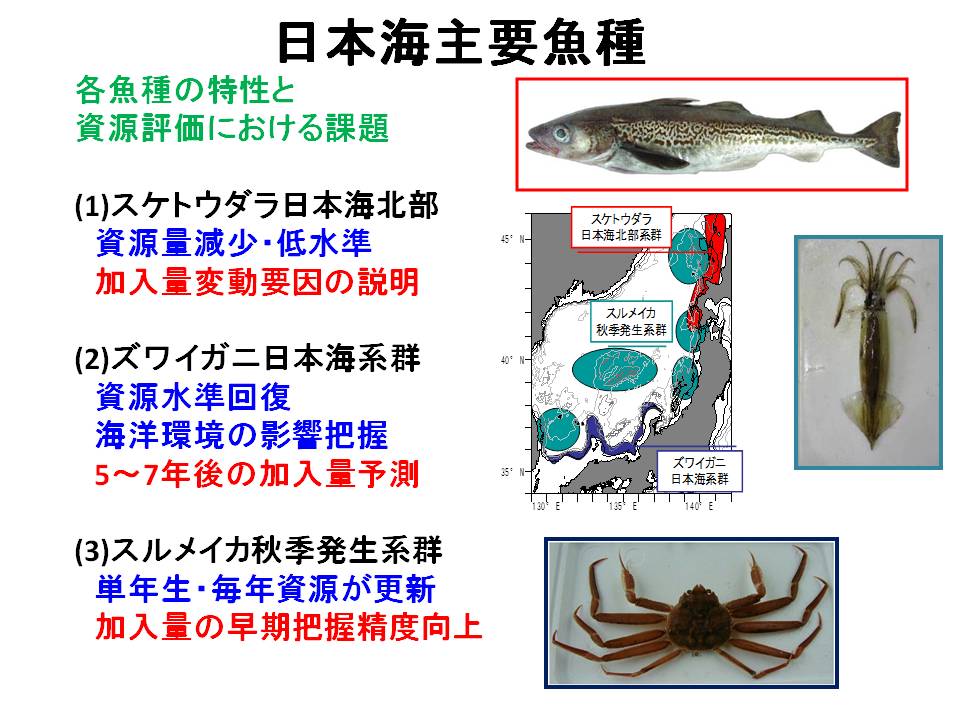
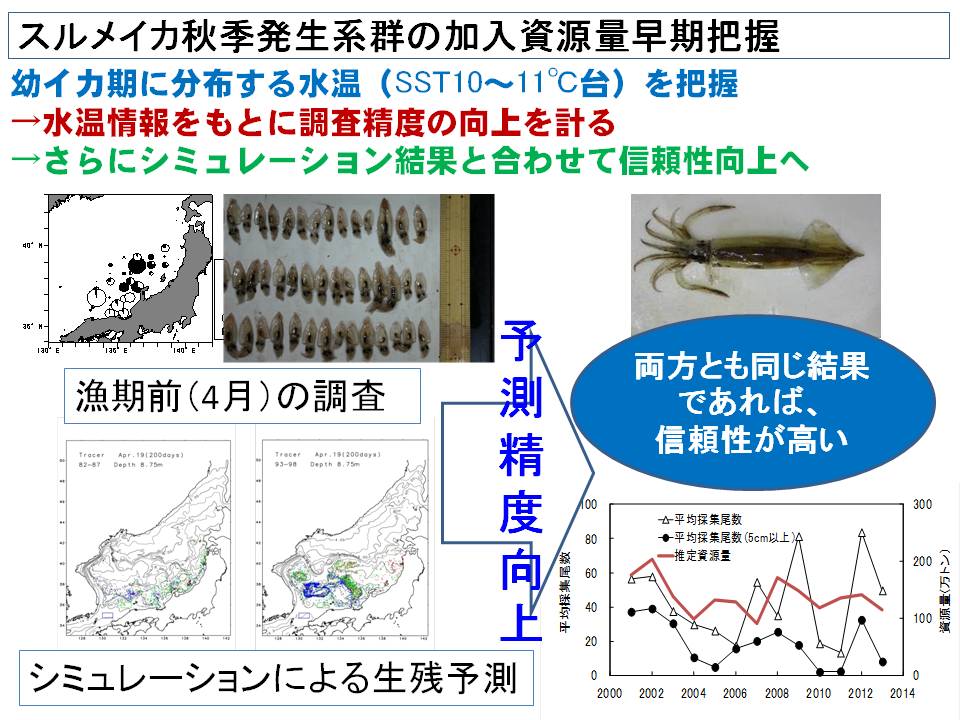
海洋環境に対する応答は、魚種・系群によって異なります。近年の日本海ではスケトウダラは資源量が減少して低位水準にあるのに対し、ズワイガニは中位水準に回復、スルメイカは中位~高位水準を維持しています。そのため、同じ日本海でも魚種系群によって直面している問題点が異なっています。例えば、スケトウダラでは低位水準を脱するための環境条件の解明、ズワイガニでは資源水準が回復した後(5~7年後)の状況予測、スルメイカでは年々の加入量の早期把握による資源管理精度の向上が求められています。
そこで本課題では、日本海の海洋環境(水温、流れ)をコンピュータ上で再現させ、各魚種系群の生残特性(水温何℃で死亡するか)とあわせて過去の海洋環境の変化に対する応答特性を明らかにすると共に、直面している問題点の解決を目指しています。
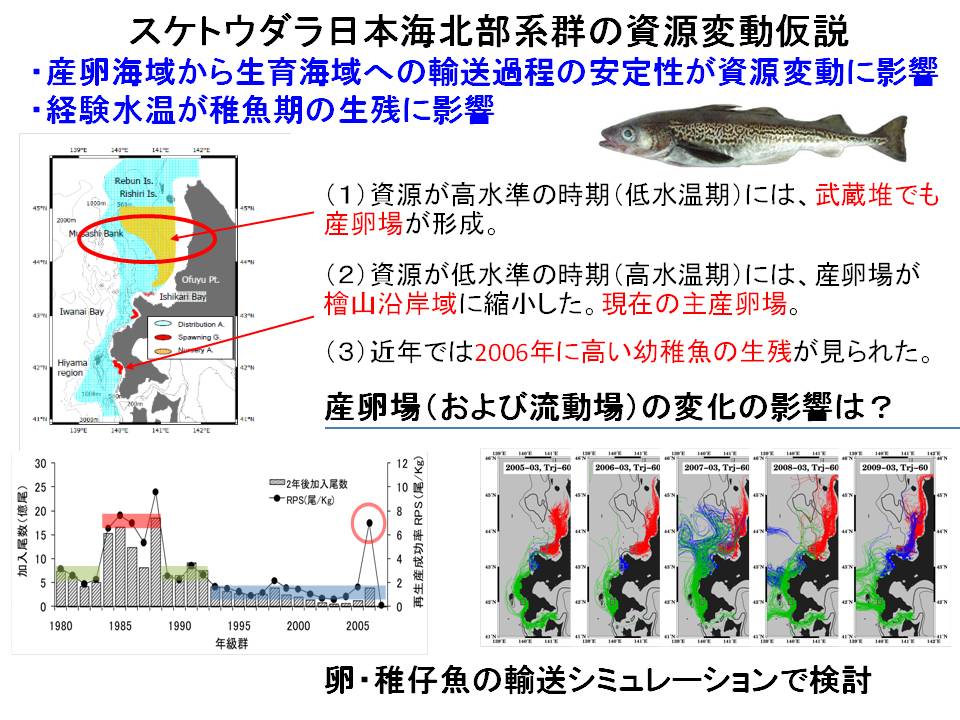
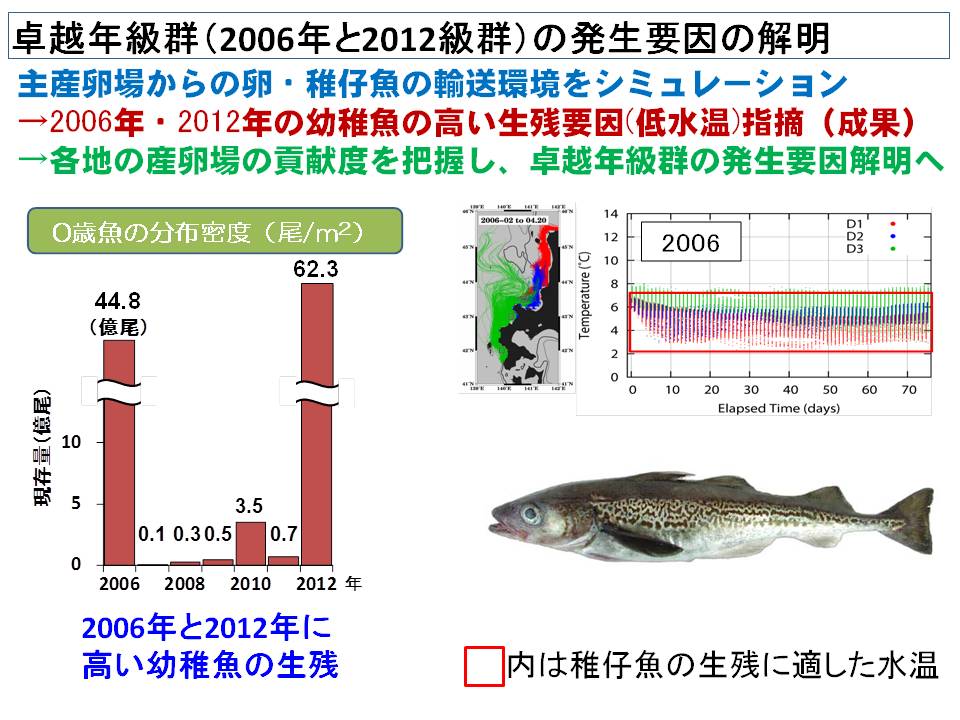
スケトウダラの資源量は卓越年級群(卵仔稚魚の生残率が高く、加入量が特に多かった年の群)の発生に大きく左右されます。スケトウダラ日本海系群では、2006年に卵仔稚魚の生残率が高く、その要因を明らかにすることが卓越年級群の発生メカニズムを解明する上で重要です。2006年の海洋環境をコンピュータ上で再現し、卵仔稚魚の生息環境を検討したところ、2006年は水温が低く、卵稚仔の生残に適していたことが示されました。さらに、2012年も卵仔稚魚の生残率が高く、2006年と同様に海洋環境をコンピュータ上で再現して要因を分析しました。その結果、2012年も水温が低く、例年よりも卵仔稚魚の生残に適していたことが示されました。つまり、海洋環境(特に低水温)が卵仔稚魚の生残を高くする条件であることが明らかとなってきました。
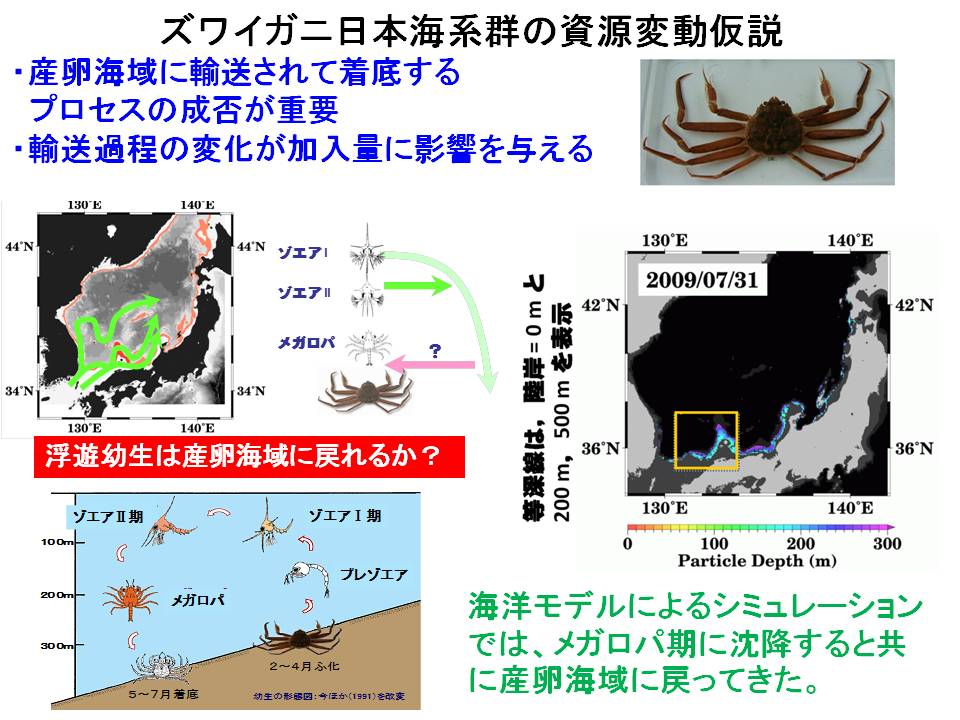
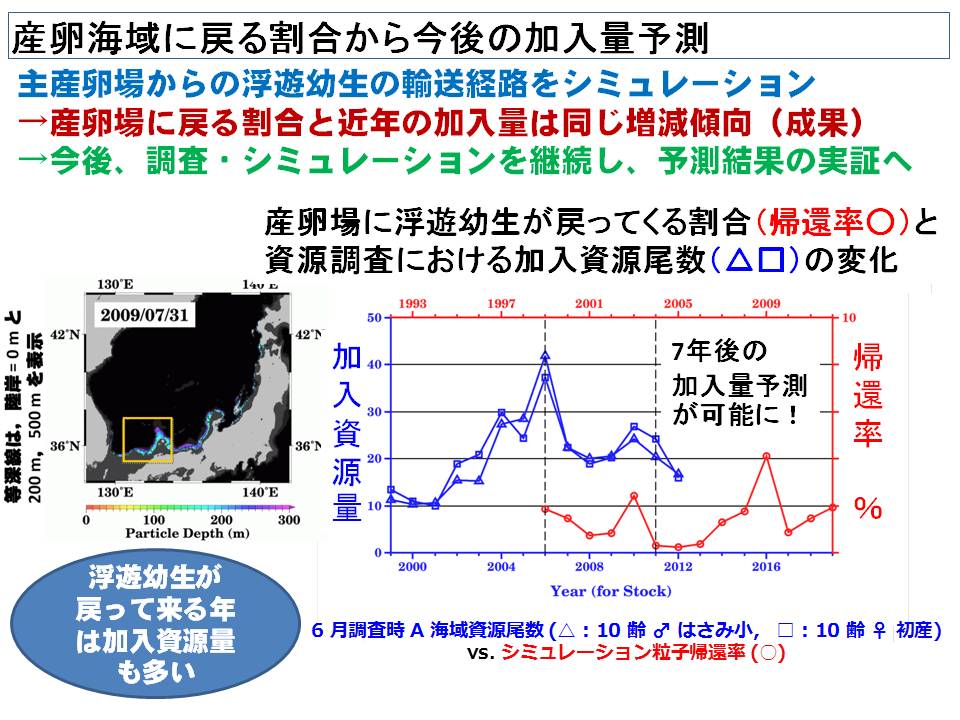
ズワイガニは底棲生物ですが、幼生はプランクトンとして、海流によって広く運ばれていきます。したがって、幼生期の拡散状況および産卵海域へどのように戻ってくるかを明らかにすることは資源変動要因を検討する際に重要です。特に、産卵海域に戻ってくる割合の変化が、資源の維持・増大とどのような関連があるのかを明らかにすることは、今後のズワイガニの資源変動を予測する上で重要です。そこで、本課題ではズワイガニ幼生の分布生態を明らかにすると共に、海洋環境をもとにシミュレーションを行いました。さらに、産卵海域へ戻ってくる幼生の割合の年変動を計算し、加入資源量(着底後7年で漁獲加入とした)との関係を検討しました。その結果、近年において、両者は同様の変動傾向であることが明らかになりました。また、今後の変化では、産卵場へ戻ってくる割合が増加する(加入資源量は増加する)と予測されました。今後の調査結果を基にこれらの予測結果を検証していく予定です。

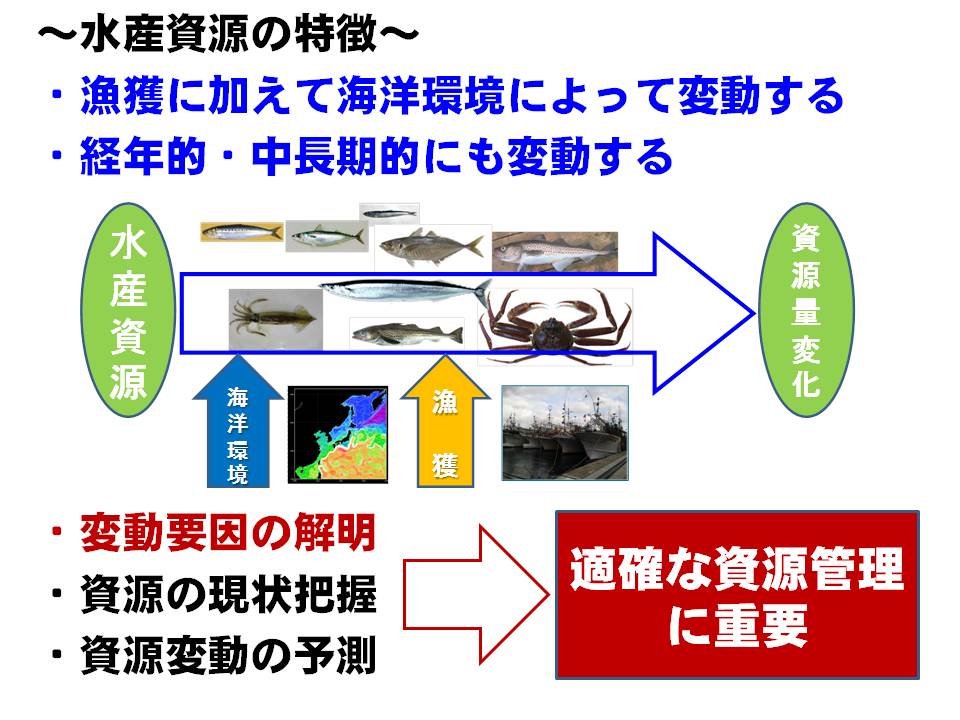
水産資源は漁業の影響だけでなく、海洋環境の影響によっても大きく変化します。しかし、私達には、海洋環境の変化をコントロールすることはほとんどできません。そのため、水産資源を適切に管理するには、海洋環境の変化に対する水産資源の応答特性を正確に把握し、資源水準と海洋環境の変化に応じた対策が重要になります。
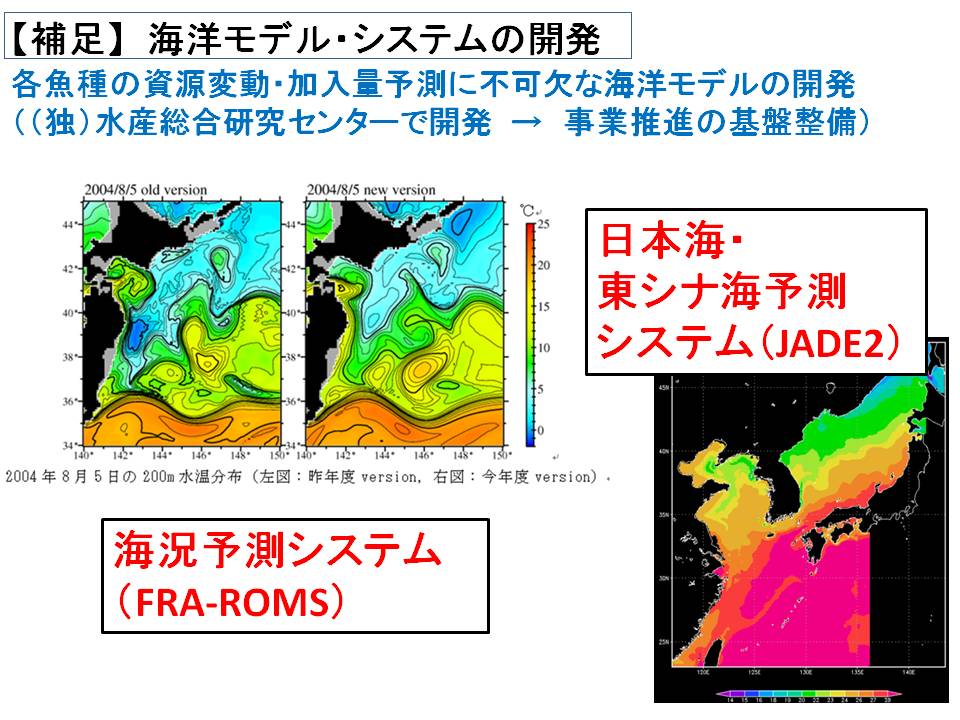
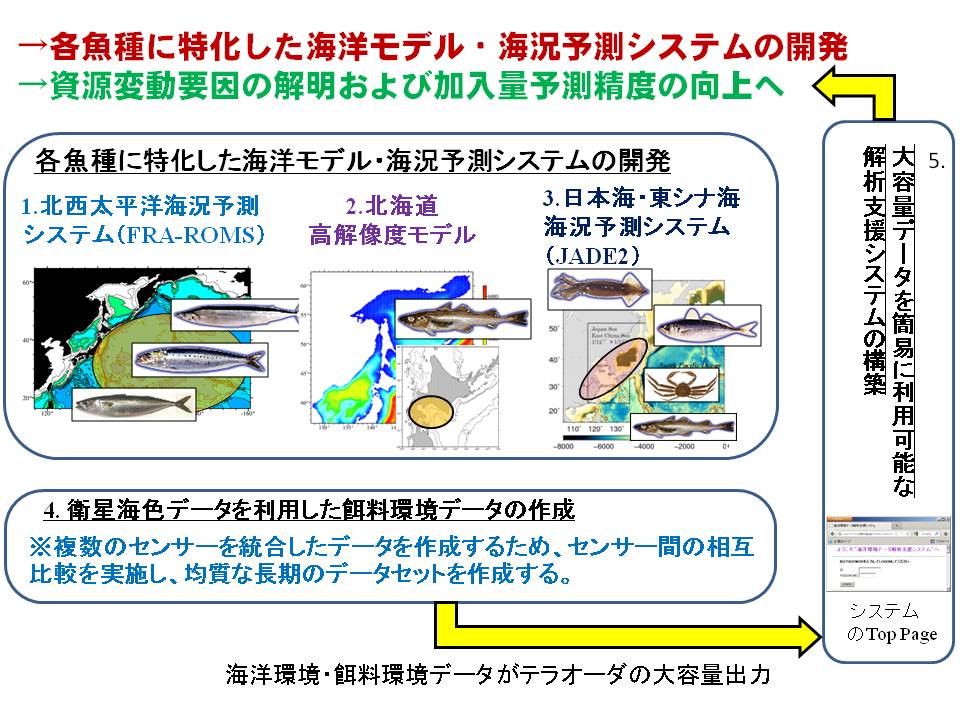
そこで本課題では、海洋環境の変化をコンピュータ上で精度良くシミュレートする海洋流動モデルを開発します(H25年度より水産総合研究センターで研究開発)。ただし、各魚種系群によって資源量の変化に影響を与える海域や海洋現象が異なるため、各魚種系群に専用の海洋流動モデルを構築し、データを作成する必要があります。このようにして作成された海洋環境データ(水温や流れ等)は各魚種系課題に提供され、資源変動機構の解明および加入量予測精度を向上させるための基礎データとして利用しています。